Glossario/Approfondimenti
E
F
G
I
M
O
P
S
T
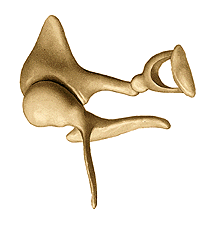 La catena degli ossicini ha lo scopo di trasferire la vibrazione della membrana timpanica alla finestra ovale determinando al tempo stesso quel processo di amplificazione della vibrazione timpanica cui si è sopra accennato.
La catena degli ossicini ha lo scopo di trasferire la vibrazione della membrana timpanica alla finestra ovale determinando al tempo stesso quel processo di amplificazione della vibrazione timpanica cui si è sopra accennato.

Il martello è, con un’estremità, a diretto contatto con il timpano; esso è poi “incernierato” all’altra estremità all’incudine, la quale, a sua volta, spinge la staffa contro la membrana della finestra ovale. È bene precisare che il processo di amplificazione non è ottenuto con il principio della leva dato che il “braccio” delle forze agenti sul martello e sull’incudine è all’incirca lo stesso; il sistema degli ossicini trasferisce praticamente inalterata (il guadagno è circa di un fattore 1.3) la forza che il timpano esercita sul martello, alla finestra ovale.
Il processo di amplificazione viene invece ottenuto applicando tale forza sulla finestra ovale la cui area è circa di un fattore 17 inferiore all’area efficace (cioè che entra realmente in vibrazione) del timpano. Infatti l'area della finestra ovale è circa 3.2 mm2, mentre l'area del timpano è circa 55 mm2.
La pressione (rapporto tra la forza normale alla superficie e la superficie stessa) è aumentata quindi circa di un fattore 22 (1.3 per gli effetti di leva meccanica e 17 per gli effetti di diminuzione dell’area).
Il processo di amplificazione della pressione (e non della forza) è utile in quanto la pressione esercitata sulla finestra ovale si trasferisce al liquido contenuto nella coclea. A causa della loro sostanziale incomprimibilità i liquidi sono in grado, secondo il principio di Pascal, di trasferire efficacemente la pressione esercitata su di una loro parte, all’intero liquido; per capire quanto è efficace la soluzione che la natura ha sviluppato per trasferire l’energia dell’onda sonora al liquido cocleare, si pensi che se fosse direttamente l’aria a mettere in moto il liquido a causa del pessimo adattamento di impedenza tra i due mezzi, (l’aria ha un impedenza acustica di 41,5 ohm acustici, il fluido cocleare di 143000) solo 1/1000 dell’energia dell’onda sonora verrebbe trasferita al liquido e il resto verrebbe riflessa impedendoci, di fatto, di sentire!
Ulteriori dettagli e approfontimenti possono essere reperiti a questo link.
Cellule Ciliate

Cellule cigliate esterne
Le cellule cigliate esterne percepiscono le vibrazioni della membrana basilare mediante i loro fasci di stereociglia. Le stereociglia si trovano immerse nell'endolinfa, un fluido che possiede un potenziale elettrico di +80 mV rispetto a quello della perilinfa che circonda la parte inferiore della cellula.
Sempre rispetto a questo, l'interno delle cellule possiede invece un potenziale negativo di -70 mV, mantenuto dalle potenti batterie a sodio e potassio delle membrane cellulari. Pertanto, attraverso le stereociglia si forma una differenza di potenziale di circa 150 mV capace di generare correnti elettriche fino a 8 nA.
Quando le stereociglia più alte si inclinano leggermente per effetto delle oscillazioni della membrana tectoria, il fascio stereocigliare si apre a ventaglio, mettendo in tensione filamenti proteici che fanno aprire i canali stereocigliari (piccole valvole poste sulle sommità delle stereociglia).
L'apertura di questi canali determina l'ingresso di correnti elettriche sufficienti a depolarizzare la cellula di alcuni millivolt, causando la contrazione della cellula.
Cellule cigliate interne
Le cellule cigliate interne sono localizzate nell'organo di Corti verso l'interno rispetto all'asse della chiocciola, queste cellule (non contrattili) sono i sensori che trasmettono i segnali al nervo acustico.
Come nel caso delle cellule cigliate esterne, l'apertura dei canali stereocigliari determina l'ingresso di una corrente elettrica che fa cadere il potenziale intracellulare. Le stereociglia più alte delle cellule cigliate interne, a differenza da quelle esterne, non sono fissate alla membrana tectoria e pertanto la loro deflessione non è proporzionale allo spostamento relativo della membrana. Esse subiscono invece l'azione viscosa del fluido messo in moto dalle oscillazioni della membrana tectoria. In questo modo la forza agente sulle stereociglia risulta proporzionale alla velocità della membrana. Questo serve a equalizzare la risposta elettrica ai segnali di alta frequenza compensando la caduta di potenziale elettrico causata dalla capacità parassita della membrana cellulare.
In condizioni di risonanza la membrana tettoriale attiva le cellule motrici con un ritardo di un quarto del periodo rispetto all'oscillazione della membrana basilare. In questo modo, le forze applicate dalle cellule motrici alla membrana basilare si comportano come le spinte che bisogna imprimere ad un'altalena per aumentarne il movimento. Anche le oscillazioni delle cellule ciliate interne seguono quelle della membrana tettoriale col ritardo di un quarto di periodo.
Da questo si deduce come le cellule cigliate siano il vero e proprio trasduttore meccanico elettrico e gli impulsi elettrici da essi generati sono inviati, attraverso i neuroni, al sistema nervoso centrale.
Ulteriori dettagli possono essere reperiti a questo link.
La parola chiralità, deriva dal greco χειρ (kheir) che significa "mano".
Essa si riferisce a molecole in cui gli atomi, o gruppi di atomi, che le costituiscono, possono essere disposti nello spazio in maniera asimmetrica. Esistono quindi coppie di molecole che impiegano gli stessi atomi ma disposti nello spazio in modo speculare. Esattamente come le due mani che non possono essere sovrapposte identicamente l'una all'altra ma possono "guardarsi allo specchio". Due molecole siffatte sono dette enantiomeri.
L'enantiomeria è un caso particolare in cui le configurazioni sono due e sono una speculare all'altra. Facendo un siscorso più generale, ci possono essere delle molecole in cui gli atomi sono legati fra loro nello stesso modo, ma con orientamenti diversi e non sempre questi orientamenti sono tali da produrre delle copie speculari.
Queste molecole sono definite stereoisomeri e le copie non sovrapponibili sono dette epimeri.
Da un punto di vista fisico, gli stereoisomeri di una molecola hanno tutti le stesse proprietà, per esempio il colore, il punto di fusione, ecc. e chimicamente reagiscono tutte nello stesso modo con molecole non chirali.
Quello che invece cambia è il modo con cui reagiscono con altre molecole chirali. Siccome fra quelle organiche, di molecole chirali ce ne sono in abbondanza, il fatto che un composto sia un epimero piuttosto che un altro fa una differenza notevole. Per esempio fra gli zuccheri, quelli importanti biologicamente sono prevalentemente quelli della serie D caratterizzati da uno specifico orientamento di uno dei gruppi H-C-OH.
Lo studioso che ha dato un contributo fondamentale in questo ambito della chimica è Emil Fischer.
Nei suoi studi sulla stereoisomeria, Fischer definì quella che poi è stata chiamata proiezione di Fischer.
Si tratta di un insieme di convenzioni per rappresentare nel piano le posizioni degli atomi nello spazio.
Nell'immagine che segue si può vedere una molecola semplificata in cui si evidenzia un centro chirale, attraverso cui si fa passare il piano di proiezione, e 4 gruppi funzionali A, B, X, e Y che sono legati al centro chirale.
Nel processo di proiezione i quattro gruppi vengono a formare una croce in cui i segmenti A e B, oltre ad essere a destra e sinistra del centro, sono anche SOPRA il piano di proiezione, mentre gli altri due sono SOTTO.
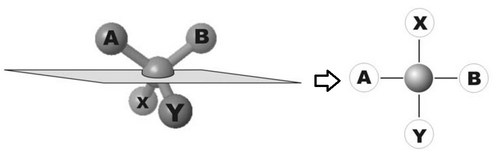
In questa rappresentazione, scambiare AB con XY significa far scavalcare tridimensionalmente il piano di proiezione, cioè si trasforma un enantiomero nell'altro.
Fischer applicò questa tecnica alla rappresentazione della gliceraldeide portando alla definizione della D-gliceraldeide e L-gliceraldeide.
Come abbiamo detto, gli zuccheri rilevanti da un punto di vista biologico sono quelli della serie "D", ma anche gli amminoacidi hanno caratteristiche stereoisomeriche e anche in questo caso, neglio organismi viventi, solo una famiglia, denominata "L" viene effettivamente utilizzata nelle cellule.
Estratto dal
PREAMBLE - WORLD HEALTH ORGANIZATION INTERNATIONAL AGENCY FOR RESEARCH ON CANCER
- Group 1: The agent is carcinogenic to humans.
This category is used when there is sufficient evidence of carcinogenicity in humans. Exceptionally, an agent may be placed in this category when evidence of carcinogenicity in humans is less than sufficient but there is sufficient evidence of carcinogenicity in experimental animals and strong evidence in exposed humans that the agent acts through a relevant mechanism of carcinogenicity. - Group 2.
This category includes agents for which, at one extreme, the degree of evidence of carcinogenicity in humans is almost sufficient, as well as those for which, at the other extreme, there are no human data but for which there is evidence of carcinogenicity in experimental animals. Agents are assigned to either Group 2A (probably carcinogenic to humans) or Group 2B (possibly carcinogenic to humans) on the basis of epidemiological and experimental evidence of carcinogenicity and mechanistic and other relevant data. The terms probably carcinogenic and possibly carcinogenic have no quantitative significance and are used simply as descriptors of different levels of evidence of human carcinogenicity, with probably carcinogenic signifying a higher level of evidence than possibly carcinogenic.- Group 2A: The agent is probably carcinogenic to humans.
This category is used when there is limited evidence of carcinogenicity in humans and sufficient evidence of carcinogenicity in experimental animals. In some cases, an agent may be classified in this category when there is inadequate evidence of carcinogenicity in humans and sufficient evidence of carcinogenicity in experimental animals and strong evidence that the carcinogenesis is mediated by a mechanism that also operates in humans. Exceptionally, an agent may be classified in this category solely on the basis of limited evidence of carcinogenicity in humans. An agent may be assigned to this category if it clearly belongs, based on mechanistic considerations, to a class of agents for which one or more members have been classified in Group 1 or Group 2A. - Group 2B: The agent is possibly carcinogenic to humans.
This category is used for agents for which there is limited evidence of carcinogenicity in humans and less than sufficient evidence of carcinogenicity in experimental animals. It may also be used when there is inadequate evidence of carcinogenicity in humans but there is sufficient evidence of carcinogenicity in experimental animals. In some instances, an agent for which there is inadequate evidence of carcinogenicity in humans and less than sufficient evidence of carcinogenicity in experimental animals together with supporting evidence from mechanistic and other relevant data may be placed in this group. An agent may be classified in this category solely on the basis of strong evidence from mechanistic and other relevant data.
- Group 2A: The agent is probably carcinogenic to humans.
- Group 3: The agent is not classifiable as to its carcinogenicity to humans.
This category is used most commonly for agents for which the evidence of carcinogenicity is inadequate in humans and inadequate or limited in experimental animals.
Exceptionally, agents for which the evidence of carcinogenicity is inadequate in humans but sufficient in experimental animals may be placed in this category when there is strong evidence that the mechanism of carcinogenicity in experimental animals does not operate in humans.
Agents that do not fall into any other group are also placed in this category.
An evaluation in Group 3 is not a determination of non-carcinogenicity or overall safety. It often means that further research is needed, especially when exposures are widespread or the cancer data are consistent with differing interpretations. - Group 4: The agent is probably not carcinogenic to humans.
This category is used for agents for which there is evidence suggesting lack of carcinogenicity in humans and in experimental animals. In some instances, agents for which there is inadequate evidence of carcinogenicity in humans but evidence suggesting lack of carcinogenicity in experimental animals, consistently and strongly supported by a broad range of mechanistic and other relevant data, may be classified in this group.
La codifica 128 non è univoca, ma, in alcuni casi, gli stessi dati possono essere rappresentati utilizzando differenti tabelle e differenti combinazioni di caratteri di start.
Quello che segue è l'algoritmo che permette di decidere quale codifica utilizzare per minimizzare la lunghezza del codice
- - Determinazione del carattere di Start:
- 1a - se i dati cominciano con 4 o piu' cifre, usare il carattere di Start C;
- 1b - altrimenti se nei dati compare un carattere di controllo prima di qualsiasi carattere alfabetico minuscolo, usare il carattere si Start A;
- 1c - altrimenti, usare il carattere di Start B.
- - Se e' verificata la condizione 1a con un numero dispari di cifre, inserire un carattere Code A o Code B prima dell'ultima cifra, seguendo le regole 1b e 1c per scegliere tra Code A e Code b.
- - Se si presentano 4 o piu' cifre numeriche mentre si e' in Code A o in Code B:
- 3a - se le cifre sono in numero pari, inserire un carattere Code C prima della prima cifra;
- 3b - vicersa, se le cifre sono in numero dispari, insrire in carattere Code C dopo la prima cifra.
- - Se si presenta un carattere di controllo mentre si e' Code B:
- 4a - se tra il carattere di controllo in questione e il successivo c'e' un carattere alfabetico minuscolo, inserire un carattere di shift prima del carattere di controllo;
- 4b - viceversa, inserire un carattere Code A prima del carattere di controllo.
- - Se compare un carattere alfabetico minuscolo mentre si e' in Code A:
- 5a - se tra il carattere alfabetico minuscolo in questione ed il successivo c'e' un carattere di controllo, inserire un carattere di shift prima del carattere alfabetico minuscolo;
- 5b - altrimenti, inserire un carattere Code B prima del carattere alfabetico minuscolo.
- - Se si ha un carattere non numerico mentre si e' in Code C, inserire un carattere Code A o Code B prima del carattere in questione, seguendo le regole 1b e 1c per scegliere tra Code A e Code B.
Tabella completa Codice 128
| Valore | Code A | Code B | Code C | B1 | S1 | B2 | S2 | B3 | S3 |
|
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 |
SP ! " # $ % & ' ( ) * + , - . / 0 1 2 3 4 5 6 7 8 9 : ; ° = > ? @ A B C D E F G H I J K L M N O P Q R S T U V W X Y Z [ \ ] ^ _ NUL SOH STX ETX EOT ENQ ACK BEL BS HT LF VT FF CR SO SI DLE DC1 DC2 DC3 DC4 NAK SYN ETB CAN EM SUB ESC FS GS RS US FNC 3 FNC 2 SHIFT Code C Code B FNC 4 FNC 1 |
SP ! " # $ % & ' ( ) * + , - . / 0 1 2 3 4 5 6 7 8 9 : ; ° = > ? @ A B C D E F G H I J K L M N O P Q R S T U V W X Y Z [ \ ] ^ _ ' a b c d e f g h i j k l m n o p q r s t u v w x y z { | } ~ DEL FNC 3 FNC 2 SHIFT Code C FNC 4 Code A FNC 1 |
00 01 02 03 04 05 06 07 08 09 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 Code B Code A FNC 1 |
2 2 2 1 1 1 1 1 1 2 2 2 1 1 1 1 1 1 2 2 2 2 2 3 3 3 3 3 3 3 2 2 2 1 1 1 1 1 1 2 2 2 1 1 1 1 1 1 3 2 2 2 2 2 3 3 3 3 3 3 3 2 4 1 1 1 1 1 1 1 1 1 1 1 1 2 2 4 2 1 1 1 1 1 1 1 4 4 4 2 2 4 1 1 1 1 1 4 4 1 1 3 4 |
1 2 2 2 2 3 2 2 3 2 2 3 1 2 2 1 2 2 2 2 2 1 2 1 1 2 2 1 2 2 1 1 3 1 3 3 1 3 3 1 3 3 1 1 3 1 1 3 1 1 3 1 1 1 1 1 3 1 1 3 1 2 3 1 1 2 2 4 4 1 1 2 2 4 4 4 2 1 4 3 1 2 2 1 2 2 1 2 2 1 1 1 1 1 3 1 1 1 1 1 1 1 1 |
2 2 2 1 1 1 2 2 2 1 1 1 2 2 2 3 3 3 3 1 1 3 3 2 1 1 1 2 2 2 2 2 2 1 1 1 2 2 2 1 1 1 2 2 2 3 3 3 3 1 1 3 3 3 1 1 1 2 2 2 4 1 1 1 1 1 1 1 1 2 2 2 2 2 2 1 1 3 1 4 1 1 1 4 4 4 1 1 1 2 4 2 1 1 1 4 4 1 1 3 4 1 1 |
2 1 2 2 3 2 2 3 2 2 3 2 2 1 2 2 1 2 2 1 2 2 1 1 2 1 2 2 1 2 1 3 1 3 1 3 3 1 3 3 1 3 1 3 1 1 3 1 1 3 1 1 3 1 1 3 1 1 3 1 1 4 1 2 4 1 4 1 2 2 4 1 4 1 2 2 1 1 1 1 2 1 2 2 1 2 2 1 2 1 1 1 1 3 1 1 3 1 3 1 1 1 1 |
2 2 2 2 2 2 1 1 1 1 1 1 3 3 3 2 2 2 1 3 3 1 1 3 2 2 2 1 1 1 2 2 2 2 2 2 1 1 1 1 1 1 3 3 3 2 2 2 2 3 3 1 1 3 2 2 2 1 1 1 1 1 1 2 2 2 2 2 2 1 1 1 1 1 1 1 1 1 1 1 4 4 4 1 1 1 1 1 1 4 2 2 4 4 4 1 1 1 1 4 3 4 3 |
2 2 1 3 2 2 3 2 2 3 2 2 2 2 1 2 2 1 1 2 1 2 2 1 2 2 1 2 2 1 3 1 1 3 3 1 3 3 1 3 3 1 3 1 1 3 1 1 1 1 1 3 1 1 3 1 1 3 1 1 1 1 1 4 2 4 1 2 1 4 2 4 1 2 1 1 4 1 2 1 2 2 1 2 2 1 2 2 1 1 1 1 3 1 1 3 1 3 1 1 1 1 1 |
| 103 104 105 |
Start A | Start B |
Start C |
2 2 2 |
1 1 1 |
1 1 1 |
4 2 2 |
1 1 3 |
2 4 2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Dove:
- B1 - B4 = barra 1 - 4
- S1 - S3 = spazio 1 - 3
- 1 = elemento di spessore 1 modulo
- 2 = elemento di spessore 2 moduli
- 3 = elemento di spessore 3 moduli
- 4 = elemento di spessore 4 moduli
Codici EAN per le prime cifre identificative del paese
| 00-13: USA & Canada | 20-29: In-Store Functions | 30-37: France |
| 40-44: Germany | 45: Japan (also 49) | 46: Russian Federation |
| 471: Taiwan | 474: Estonia | 475: Latvia |
| 477: Lithuania | 479: Sri Lanka | 480: Philippines |
| 482: Ukraine | 484: Moldova | 485: Armenia |
| 486: Georgia | 487: Kazakhstan | 489: Hong Kong |
| 49: Japan (JAN-13) | 50: United Kingdom | 520: Greece |
| 528: Lebanon | 529: Cyprus | 531: Macedonia |
| 535: Malta | 539: Ireland | 54: Belgium & Luxembourg |
| 560: Portugal | 569: Iceland | 57: Denmark |
| 590: Poland | 594: Romania | 599: Hungary |
| 600 & 601: South Africa | 609: Mauritius | 611: Morocco |
| 613: Algeria | 619: Tunisia | 622: Egypt |
| 625: Jordan | 626: Iran | 64: Finland |
| 690-692: China | 70: Norway | 729: Israel |
| 73: Sweden | 740: Guatemala | 741: El Salvador |
| 742: Honduras | 743: Nicaragua | 744: Costa Rica |
| 746: Dominican Republic | 750: Mexico | 759: Venezuela |
| 76: Switzerland | 770: Colombia | 773: Uruguay |
| 775: Peru | 777: Bolivia | 779: Argentina |
| 780: Chile | 784: Paraguay | 785: Peru |
| 786: Ecuador | 789: Brazil | 80 - 83: Italy |
| 84: Spain | 850: Cuba | 858: Slovakia |
| 859: Czech Republic | 860: Yugloslavia | 869: Turkey |
| 87: Netherlands | 880: South Korea | 885: Thailand |
| 888: Singapore | 890: India | 893: Vietnam |
| 899: Indonesia | 90 & 91: Austria | 93: Australia |
| 94: New Zealand | 955: Malaysia | 977: International Standard Serial Number for Periodicals (ISSN) |
| 978: International Standard Book Numbering (ISBN) | 979: International Standard Music Number (ISMN) | 980: Refund receipts |
| 981 & 982: Common Currency Coupons | 99: Coupons | |
| Dec. | Binary | EFM |
|---|---|---|
|
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 145 146 147 148 149 150 151 152 153 154 155 156 157 158 159 160 161 162 163 164 165 166 167 168 169 170 171 172 173 174 175 176 177 178 179 180 181 182 183 184 185 186 187 188 189 190 191 192 193 194 195 196 197 198 199 200 201 202 203 204 205 206 207 208 209 210 211 212 213 214 215 216 217 218 219 220 221 222 223 224 225 226 227 228 229 230 231 232 233 234 235 236 237 238 239 240 241 242 243 244 245 246 247 248 249 250 251 252 253 254 255 |
00000000 00000001 00000010 00000011 00000100 00000101 00000110 00000111 00001000 00001001 00001010 00001011 00001100 00001101 00001110 00001111 00010000 00010001 00010010 00010011 00010100 00010101 00010110 00010111 00011000 00011001 00011010 00011011 00011100 00011101 00011110 00011111 00100000 00100001 00100010 00100011 00100100 00100101 00100110 00100111 00101000 00101001 00101010 00101011 00101100 00101101 00101110 00101111 00110000 00110001 00110010 00110011 00110100 00110101 00110110 00110111 00111000 00111001 00111010 00111011 00111100 00111101 00111110 00111111 01000000 01000001 01000010 01000011 01000100 01000101 01000110 01000111 01001000 01001001 01001010 01001011 01001100 01001101 01001110 01001111 01010000 01010001 01010010 01010011 01010100 01010101 01010110 01010111 01011000 01011001 01011010 01011011 01011100 01011101 01011110 01011111 01100000 01100001 01100010 01100011 01100100 01100101 01100110 01100111 01101000 01101001 01101010 01101011 01101100 01101101 01101110 01101111 01110000 01110001 01110010 01110011 01110100 01110101 01110110 01110111 01111000 01111001 01111010 01111011 01111100 01111101 01111110 01111111 10000000 10000001 10000010 10000011 10000100 10000101 10000110 10000111 10001000 10001001 10001010 10001011 10001100 10001101 10001110 10001111 10010000 10010001 10010010 10010011 10010100 10010101 10010110 10010111 10011000 10011001 10011010 10011011 10011100 10011101 10011110 10011111 10100000 10100001 10100010 10100011 10100100 10100101 10100110 10100111 10101000 10101001 10101010 10101011 10101100 10101101 10101110 10101111 10110000 10110001 10110010 10110011 10110100 10110101 10110110 10110111 10111000 10111001 10111010 10111011 10111100 10111101 10111110 10111111 11000000 11000001 11000010 11000011 11000100 11000101 11000110 11000111 11001000 11001001 11001010 11001011 11001100 11001101 11001110 11001111 11010000 11010001 11010010 11010011 11010100 11010101 11010110 11010111 11011000 11011001 11011010 11011011 11011100 11011101 11011110 11011111 11100000 11100001 11100010 11100011 11100100 11100101 11100110 11100111 11101000 11101001 11101010 11101011 11101100 11101101 11101110 11101111 11110000 11110001 11110010 11110011 11110100 11110101 11110110 11110111 11111000 11111001 11111010 11111011 11111100 11111101 11111110 11111111 |
01001000100000 10000100000000 10010000100000 10001000100000 01000100000000 00000100010000 00010000100000 00100100000000 01001001000000 10000001000000 10010001000000 10001001000000 01000001000000 00000001000000 00010001000000 00100001000000 10000000100000 10000010000000 10010010000000 00100000100000 01000010000000 00000010000000 00010010000000 00100010000000 01001000010000 10000000010000 10010000010000 10001000010000 01000000010000 00001000010000 00010000010000 00100000010000 00000000100000 10000100001000 00001000100000 00100100100000 01000100001000 00000100001000 01000000100000 00100100001000 01001001001000 10000001001000 10010001001000 10001001001000 01000001001000 00000001001000 00010001001000 00100001001000 00000100000000 10000010001000 10010010001000 10000100010000 01000010001000 00000010001000 00010010001000 00100010001000 01001000001000 10000000001000 10010000001000 10001000001000 01000000001000 00001000001000 00010000001000 00100000001000 01001000100100 10000100100100 10010000100100 10001000100100 01000100100100 00000000100100 00010000100100 00100100100100 01001001000100 10000001000100 10010001000100 10001001000100 01000001000100 00000001000100 00010001000100 00100001000100 10000000100100 10000010000100 10010010000100 00100000100100 01000010000100 00000010000100 00010010000100 00100010000100 01001000000100 10000000000100 10010000000100 10001000000100 01000000000100 00001000000100 00010000000100 00100000000100 01001000100010 10000100100010 10010000100010 10001000100010 01000100100010 00000000100010 01000000100100 00100100100010 01001001000010 10000001000010 10010001000010 10001001000010 01000001000010 00000001000010 00010001000010 00100001000010 10000000100010 10000010000010 10010010000010 00100000100010 01000010000010 00000010000010 00010010000010 00100010000010 01001000000010 00001001001000 10010000000010 10001000000010 01000000000010 00001000000010 00010000000010 00100000000010 01001000100001 10000100100001 10010000100001 10001000100001 01000100100001 00000000100001 00010000100001 00100100100001 01001001000001 10000001000001 10010001000001 10001001000001 01000001000001 00000001000001 00010001000001 00100001000001 10000000100001 10000010000001 10010010000001 00100000100001 01000010000001 00000010000001 00010010000001 00100010000001 01001000000001 10000010010000 10010000000001 10001000000001 01000010010000 00001000000001 00010000000001 00100010010000 00001000100001 10000100001001 01000100010000 00000100100001 01000100001001 00000100001001 01000000100001 00100100001001 01001001001001 10000001001001 10010001001001 10001001001001 01000001001001 00000001001001 00010001001001 00100001001001 00000100100000 10000010001001 10010010001001 00100100010000 01000010001001 00000010001001 00010010001001 00100010001001 01001000001001 10000000001001 10010000001001 10001000001001 01000000001001 00001000001001 00010000001001 00100000001001 01000100100000 10000100010001 10010010010000 00001000100100 01000100010001 00000100010001 00010010010000 00100100010001 00001001000001 10000100000001 00001001000100 00001001000000 01000100000001 00000100000001 00000010010000 00100100000001 00000100100100 10000010010001 10010010010001 10000100100000 01000010010001 00000010010001 00010010010001 00100010010001 01001000010001 10000000010001 10010000010001 10001000010001 01000000010001 00001000010001 00010000010001 00100000010001 01000100000010 00000100000010 10000100010010 00100100000010 01000100010010 00000100010010 01000000100010 00100100010010 10000100000010 10000100000100 00001001001001 00001001000010 01000100000100 00000100000100 00010000100010 00100100000100 00000100100010 10000010010010 10010010010010 00001000100010 01000010010010 00000010010010 00010010010010 00100010010010 01001000010010 10000000010010 10010000010010 10001000010010 01000000010010 00001000010010 00010000010010 00100000010010 |
Il sistema acustico utilizza un secondo sistema per codificare le varie frequenze oltre alla codifica tonotopica della coclea.
Fino a frequenze nell'ordine dei 3kHz la coclea può utilizzare direttamente un sistema di aggancio di fase in cui un singolo neurone può scaricare in un determinato punto dell'onda sonora ad una certa frequenza. I pattern di scarica determinano quindi un codice temporale.

Per frequenze più alte, fino a 5kHz, è stato proposto il principio di scarica tramite il quale è necessaria la collaborazione di più neuroni per ricostruire il pattern.

Oltre i 5kHz la velocità di scarica dei neuroni è comunque troppo lenta, quindi per i suoni più acuti il cervello si deve basare solo su informazioni derivanti dalla zona di membrana oggetto dell'eccitazione.
I conettivi logici sono utilizzati sia nell'ambito del linguaggio naturale che nell'ambito della logica formale per creare dei collegamenti fra proposizioni al fine di ottenere una proposizione risultante il cui valore di verità sia opportunamente ricavato dalle proposizioni originali.
Normalmente si utilizzano le lettere A, B e C per indicare le due proposizioni di partenza e la terza proposizione risultante e le lettere V e F per indicare il valore di verità delle stesse.
Siccome le operazioni logiche possono essere facilmente implementate all'interno dei dispositivi elettronici, sia come veri e propri circuiti hardware, sia come operazioni svolte dal software, è frequente utilizzare i simboli 0 e 1 per identificare i livelli di verità.
I connettivi logici si dividono in binari, se accettano come argomento due proposizioni, e unari se ne accettano solo una.
Normalmente la definizione dei connettivi logici avviene attraverso le cosiddette tabelle di verità. Cioè delle tabelle che riportano il valore di verità della proposizione risultante C per ogni combinazione di valori che possono assumere le due proposizioni A e B in ingresso.
Elenchiamo di seguito i principali connettivi logici.
Congiunzione logica
La congiunzione logica può essere indicata come, a seconda dei contesti:
e nel linguaggio comune, et (dal latino) nella logica fromale, AND in logica booleana, con il simbolo ∧
La congiunzione logica assume valore vero quando entrambe le proposizioni in ingresso hanno valore vero.
| A | B | C=A AND B |
|---|---|---|
| 0 | 0 | 0 |
| 0 | 1 | 0 |
| 1 | 0 | 0 |
| 1 | 1 | 1 |
Disgiunzione Inclusiva
La disgiunzione inclusiva può essere indicata come, a seconda dei contesti:
o nel linguaggio comune, vel (dal latino) nella logica formale, OR in logica booleana, con il simbolo ∨
La disgiunzione inclusiva ha valore vero quando almeno una delle proposizioni in ingresso ha valore vero
| A | B | C=A OR B |
|---|---|---|
| 0 | 0 | 0 |
| 0 | 1 | 1 |
| 1 | 0 | 1 |
| 1 | 1 | 1 |
Disgiunzione Esclusiva
La disgiunzione esclusiva può essere indicata come, a seconda dei contesti:
o nel linguaggio comune, aut (dal latino) nella logica formale, XOR oppure EXOR in logica booleana, indicata dal simbolo ⊕
La disgiunzione esclusiva ha valore vero quando solo una delle proposizione in ingresso ha valore vero
| A | B | C=A XOR B |
|---|---|---|
| 0 | 0 | 0 |
| 0 | 1 | 1 |
| 1 | 0 | 1 |
| 1 | 1 | 0 |
Negazione
La negazione logica è un operatore unario che inverte il valore di verità della proposizione cui è applicato
Spesso si annovera inoltre fra i connettivi logici la negazione logica "non", indicata con il simbolo la quale agisce però su un'unica proposizione. Essa si indica con:
non nel linguaggio comune, NOT in logica booleana, con il simbolo ¬
| A | C=NOT A |
|---|---|
| 0 | 1 |
| 1 | 0 |
Gli elettroliti sono degli ioni disciolti nel sangue.
Si tratta cioè di sostanze dotate di carica delle quali deve esserci una concentrazione costante affinchè il bilancio idrico e la pressione osmotica restino nella norma ed affinchè determinati enzimi possano funzionare regolarmente.
Essi i principali sono:
- Calcio - Ca++: importante per la formazione di denti e ossa; aiuta la coagulazione, il movimento muscolare, le difese immunitarie e contribuisce alla conduzione del sistema nervoso.
- Potassio - K+: aiuta a mantenere l'equilibrio acido-base nelle cellule e la ritenzione idrica; serve all'attività muscolare, alla trasmissione nervosa e al funzionamento di determinati enzimi.
- Sodio - Na+: aiuta a mantenere l'equilibrio idrico all'interno delle cellule, oltre che contribuire al movimento muscolare e alla trasmissione nervosa. Viene assimilato tramite carne e sale.
- Cloro - Cl-: aiuta a mantenere l'equilibrio idrico nelle cellule.
La parola eterotrofia deriva dai due termini greci ἕτερος (éteros - diverso) e τροφή (trofé - nutrimento) e descrive la condizione in cui si trovano gli organismi viventi che non sono in grado di sintetizzare il proprio nutrimento autonomamente a partire da sostanze inorganiche.
Questi organismi hanno bisogno di attingere nell'ambiente i vari composti organici pre-sintetizzati da altri organismi, che sono invece detti autotrofi (ad es. tutte le piante che posseggono clorofilla).
Sono eterotrofi tutti gli animali (pluricellulari eterotrofi), i protozoi, i funghi e quasi tutti i batteri.
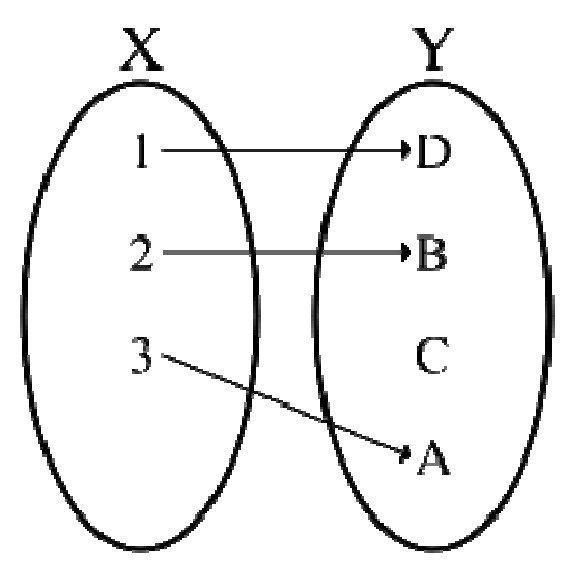 Una funzione si dice iniettiva se elementi distinti del dominio hanno un'immagine distinta, o equivalentemente se ogni elemento del codominio corrisponde al più ad un elemento del dominio.
Una funzione si dice iniettiva se elementi distinti del dominio hanno un'immagine distinta, o equivalentemente se ogni elemento del codominio corrisponde al più ad un elemento del dominio.Formalmente:
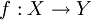 è iniettiva se
è iniettiva se 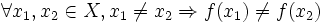
o equivalentemente:è iniettiva se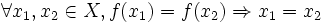
La gliceraldeide è un carboidrato aldo-trioso, ovvero costituito da tre atomi di carbonio e contenente un gruppo aldeidico (CHO).
Il nome deriva dal fatto che la molecola della gliceraldeide è simile a quella del glicerolo la cui formula è
CH2OH-CHOH-CH2OH (glicerolo)
in cui uno dei due gruppi CH2OH è stato sostituito da un gruppo CHO ottenendo
CH2OH-CHOH-CHO (gliceraldeide)
Una caratteristica importante della gliceraldeide è la sua chiralità, cioè il fatto che la sua molecola possa esistere in due configurazioni spaziali speculari.
L'asimmetria spaziale si trasforma nella proprietà di polarizzare la luce che la attraversa in un senso piuttosto che nell'altro, cioè verso destra o verso sinistra.
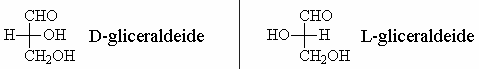
Per questo motivo le due configurazioni sono denominate D e L e sono convenzionalmente rappresentate come in figura.
La D-gliceraldeide è presa a modello per la nomenclatura dei carboidrati.
Sono detti carboidrati della serie D quelli la cui disposizione dei sostituenti attorno al penultimo atomo di carbonio è simile a quella della D-glicerladeide. Quelli a configurazione speculare nell'ultimo carbonio chirale sono invece carboidrati della serie L.
Questa classificazione e nomenclatura si deve al lavoro del chimico tedesco premio nobel per la chimica nel 1902 Hermann Emil Fischer proprio per i suoi studi sulla chimica degli zuccheri.
Fischer poteva constatare gli effetti della chiralità delle molecole degli zuccheri sulla polarizzazione della luce, quindi aveva ben chiaro che la disposizione spaziale degli atomi nelle varie molecole doveva essere diversa ma con gli strumenti a sua disposizione, non era in grado di distinguere fra zuccheri enantiomerici.
Egli introdusse allora la convenzione secondo cui gli stereoisomeri (+) e (-) della gliceraldeide venivano designati rispettivamente come:
- D-gliceraldeide -> lo stereoisomero con l’ossidrile sul C2 a destra
- L-gliceraldeide -> lo stereoisomero con l’ossidrile sul C2 a sinistra
ben sapendo che c’era soltanto il 50% delle possibilità che questa assegnazione fosse corretta.
Fischer assunse quindi che la configurazione di queste molecole fosse quella riportata in figura.
Più tardi, nel 1954, con l’avvento della diffrazione ai raggi X, Bijvoet determinò la struttura dell’acido D-tartarico determinandone così la configurazione assoluta e constatando che l’assegnazione di Fischer (ossidrile a destra nella (+)-gliceraldeide) era corretta.
Il termine deriva dalla fusione di due parole del greco antico, γλυκύς (glykýs - dolce) e λύσις (lýsis - scissione).
Si tratta infatti di un processo metabolico mediante il quale una molecola di glucosio viene scissa in due molecole di piruvato al fine di generare molecole a più alta energia, come 2 molecole di ATP e 2 molecole di NADH per ogni molecola di glucosio utilizzata.
La glicolisi avviene nel citoplasma cellulare ed è il mezzo per ottenere energia più sfruttato in natura, soprattutto grazie alla sua anaerobioticità. Probabilmente esso si sviluppò con i primi procarioti circa 3,5 miliardi di anni fa.
Il processo completo consta di dieci fasi, ciascuna associata ad un particolare enzima. In una prima fase del processo, composta da cinque passaggi, viene consumata energia per ottenere dal glucosio molecole di un derivato del glucosio a più alta energia (gliceraldeide-3-fosfato).
La gliceraldeide-3-fosfato viene poi trasformata nella fase successiva, composta di altri cinque passaggi, in molecole nettamente meno energetiche di piruvato, con produzione di energia superiore a quella consumata nella prima fase. Il processo nel suo insieme è quindi di tipo catabolico, cioè in cui molecole più complesse ed energetiche vengono trasformate in altre più semplici e meno energetiche, con accumulo di energia.
La reazione finale della glicolisi è quindi:
glucosio + 2 NAD+ + 2 ADP + 2 Pi → 2 NADH + 2 piruvato + 2 ATP + 2 H2O + 2 H+
La produzione finale del piruvato è necessaria per il ciclo di Krebs che, in accoppiata alla fosforilazione ossidativa, grazie al consumo di ossigeno si produrranno altre molecole di ATP.
Come si può vedere dalla reazione totale riportata sopra, la glicolisi ha bisogno di 2 molecole di NAD+ e produce 2 molecole di NADH. E' necessario quindi che vi sia un meccanismo metabolico che permetta al NADH di ritrasformarsi in NAD+ per ricominciare il ciclo. Negli organismi che sono in grado di effettuare un metabolismo aerobico, questo compito è devoluto al complesso della respirazione cellulare. Mentre negli organismi anaerobici, vi sono sostanzialmente due strade:
piruvato + NADH + H+ → lattato + NAD+ (fermentazione lattica)
piruvato + NADH + H+ → etanolo + CO2 + NAD+ (fermentazione alcolica)
Le varie fasi della glicolisi sono descritte in moltissimi siti su internet, uno fra tutti Wikipedia, cui si rimanda per avere ulteriori dettagli.
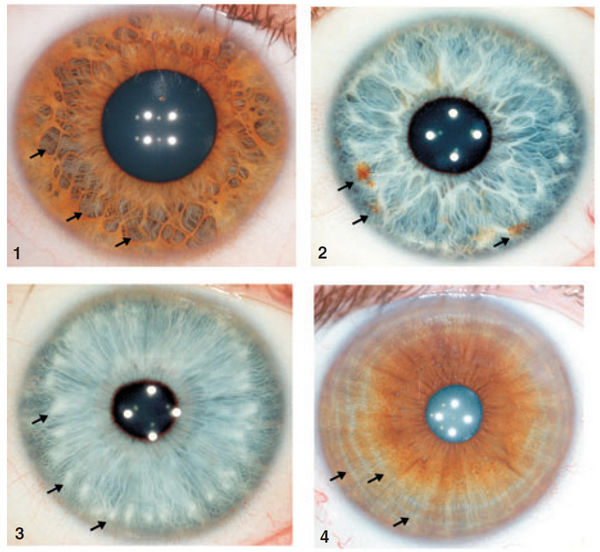
Nella foto che segue possiamo vedere tre macro-classi in cui si possono dividere i colori dell'iride nel genere umano: blu, verde/nocciola e marrone; a sinistra vediamo il caso senza anello peripupillare e a destra con anello peripupillare.

Nell'iride possono trovarsi anche delle irregolarità, difetti e disuniformità del colore legate a vari fattori.
Nell'esempio mostrato sotto sono evidenziate: 1. Cripte di Fuchs, cioè sone di moderata atrofia dello stroma; 2. Nevi, cioè punti di accumulo della melanina; 3. Noduli di Wolfflin, punti di accumulo di fibrille di collagene; 4. Pliche circolari, sottili solchi concentrici alla pupilla, i quali corrispondono a variazioni di spessore dell'epitelio pigmentato.
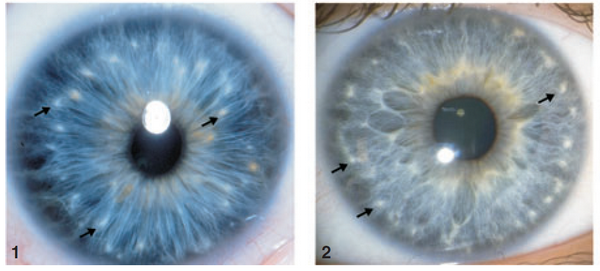
Infine, nell'immagine seguente sono messe a confronto due formazioni dovute ad accumuli puntiformi di tessuto connettivo: 1. Macchie di Brushfield, presenti nei bambini affetti da sindrome di Down i quali hanno il 35% di svilupparle; 2. Noduli di Wolfflin, osservati in soggetti normali.
La maggioranza in una votazione si distingue in:
- Assoluta: 50% + 1 degli aventi diritto
- Qualificata: 66,7% degli aventi diritto
- Relativa: maggior numero di voti
- Semplice: 50% + 1 dei votanti
L’orecchio esterno è costituito dal padiglione auricolare e dal condotto uditivo esterno, un sottile “tubo” che termina in una membrana detta timpano.
Le funzioni del padiglione auricolare sono di raccogliere una porzione significativa delle onde acustiche (proporzionale all’area del padiglione) e di convogliarle, tramite il condotto uditivo, alla membrana timpanica; determinare la localizzazione della sorgente sonora (operazione che non potrebbe essere effettuata con uguale precisione se avessimo un solo orecchio anziché due); di protezione della membrana timpanica da lesioni meccaniche; di mantenere di tale delicata membrana a condizioni di temperatura, umidità e lubrificazione costanti, in modo da preservarne le caratteristiche elastiche.
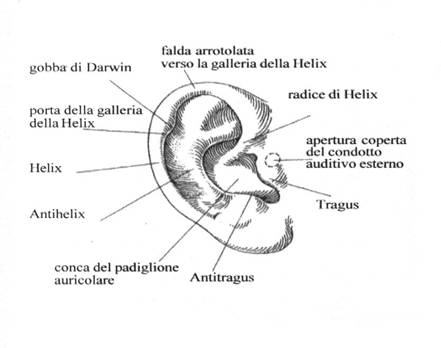
Grazie alla sofisticata conformazione delle pieghe del padiglione auricolare, il nostro cervello è in grado di misurare le differenze di fase fra i segnali che giungono alle due orecchie. Questa informazione, insieme alla differente intensità percepita, viene utilizzata per determinare la localizzazione spaziale della sorgente, sia nella direzione destra-sinistra, che alto-basso, come schematizzato nelle immagini seguenti.
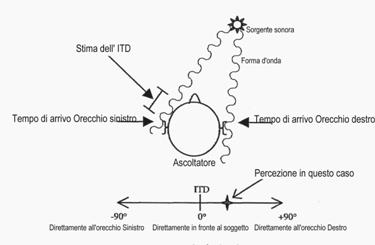

Ulteriori dettagli e approfontimenti possono essere reperiti a questo link. e a questo link da cui sono tratte anche le immagini.
Il condotto uditivo esterno ha mediamente un diametro di 7.5 mm e una lunghezza di 22-25 mm e non sembra avere altra funzione se non quella di convogliare l’onda sonora verso la membrana timpanica. In realtà la lunghezza di tale condotto gioca un ruolo decisivo nel determinare l'intervallo di frequenze di massima sensibilità uditiva.
Il condotto è in pratica un risuonatore che entra in risonanza a delle frequenze che sono tipiche della nostra specie, per le quali l'evoluzione ha selezionato la massima sensibilità. Si tratta infatti della porzione di spettro dove risiede la voce umana
Il condotto uditivo può essere modellato come un tubo chiuso la cui frequenza di risonanza corrispone a 4 volte la lunghezza dello stesso.
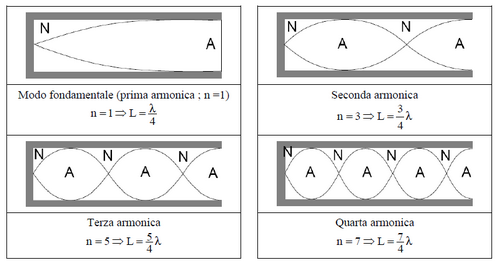
Con un rapido calcolo è facile mostrare che se si tratta il condotto come un tubo con una estremità aperta esso entra in risonanza alla frequenza di circa 2700 Hz
Nell'insieme si hanno gli effetti di diffrazione del padiglione auricolare e la forma tutt'altro che perfettamente cilindrica e rettilinea del condotto. Ne risulta che la regione di massima sensibilità uditiva si colloca attorno ai 3800 Hz.
L'organo del Corti, così chiamato in onore di Alfonso Corti che lo scoprì e lo studiò nel 1847, è una struttura cellulare, ancorata alla membrana basilare, formata da una lunga schiera di moduli affiancati tutti simili fra loro (la coclea umana ne contiene circa 3500).
Ogni modulo contiene due tipi di cellule: cellule sensoriali e cellule di supporto.

Le cellule sensoriali sono anch'esse di due tipi: cellule ciliate interne (IHC, inner hair cells) e cellule ciliate esterne (OHC, outer hair cells ); in ogni sezione radiale di coclea vi sono tre file di cellule ciliate esterne ed una fila di cellule ciliate interne.
Ogni cellula sensoriale è in contatto con diversi tipi di cellule di supporto: le cellule pillari interne ed esterne separano le OHC dalle IHC, le cellule di Deiters collegano le cellule ciliate esterne alla membrana basilare.
Lo studio moderno del fenomeno delle otoemissioni si deve al lavoro di David T. Kemp del 1978, il quale ha effettuato delle registrazioni dei suoni emessi attivamente dalla coclea umana in seguito a una stimolazione sonora.
Questi suoni si presentano con una intensità particolarmente bassa e richiedono perciò tecniche particolari per la loro evidenziazione. Nonostante alcune incertezze riguardo ai meccanismi fisiologici e al tipo di strutture cocleari coinvolte, è unanimemente accettato che le otoemissioni sono generate dalle attività contrattile delle cellule ciliate esterne e dalle caratteristiche meccano-strutturali della membrana basilare con trasformazioni di energia meccanica in energia acustica.
Le otoemissioni acustiche sono presenti sia spontaneamente (SOAE: Spontaneus Otoacustic Emissions) che in seguito a una stimolazione sonora (click o tone bursts) detto anche EOAE (Evokated Otoacustic Emissions).
Il metodo per rilevare le otoemissioni acustiche evocate, che sono le più usate nella diagnostica infantile, consiste nell’inviare degli stimoli impulsivi attraverso un’apposita sonda applicata nel condotto uditivo esterno, all’interno del quale è posto anche un microfono miniaturizzato e un tubicino di scappamento per evitare un eccessivo accoppiamento acustico tra microfono e altoparlante. Le otoemissioni così captate dal microfono vengono inviate a una apparecchiatura computerizzata per la loro analisi.
Il processo Leblanc è procedimento industriale che permette di produrre il carbonato di sodio (soda) a partire da una soluzione salina di cloruro di sodio (sale marino) e carbonato di calcio (calcare) mediante l'utilizzo di acido solforico e carbone.
Esso fu brevettato nel settembre del 1790 dal medico francese Nicolas Leblanc e conobbe subito una rapida diffusione specialmente in Francia e Inghilterra.
Il processo prevede due step successivi di reazioni chimiche:
nel primo step il cloruro di sodio viene convertito in solfato di sodio
![]()
nel secondo step, aggiungendo il carbonato di calcio e il carbone, si ottiene il carbonato di sodio.
![]()
A livello industriale, come si può vedere dalla prima reazione, fra i prodotti troviamo il solfato di sodio che rientra nel secondo step ma anche una notevole quantità di acido cloridrico gassoso. Nei primi decenni di attività delle fabbriche Leblanc l’acido cloridrico veniva disperso direttamente nell’aria.
In alternativa, specie in Francia, gli scarichi subivano un lavaggio in giare di grès piene d’acqua collegate in serie, oppure venivano convogliati in una grande camera di piombo dove reagivano con vapori ammoniacali prodotti dalla calcinazione di scarti animali, dando luogo alla formazione di cloruro d’ammonio. Si è valutato che negli anni ’40 del XIX secolo, in Inghilterra, nell’area del Merseyside, circa 100 mila tonnellate di sale siano state convertite in alcali, dando come sottoprodotto 60 mila tonnellate di acido.
 Gli addetti alla produzione che, prima dell’introduzione dei forni meccanici (mostrati nell'immagine), mescolavano manualmente l’acido solforico con il sale in grandi vasche di ghisa, si ammalavano rapidamente.
Gli addetti alla produzione che, prima dell’introduzione dei forni meccanici (mostrati nell'immagine), mescolavano manualmente l’acido solforico con il sale in grandi vasche di ghisa, si ammalavano rapidamente.
A causa del gas il bestiame soffocava, le popolazioni circostanti soffrivano di vari disturbi, i manufatti metallici e i materiali da costruzione venivano corrosi, la vegetazione e le colture agricole deperivano, i fiumi diventavano “neri come l’inchiostro” e le piogge acide completavano
l’opera di devastazione.
Inoltre il solfuro di calcio, anch'esso nocivo, veniva accumulato senza protezione alcuna all’esterno delle fabbriche, cosicché per azione delle piogge acide produceva idrogeno solforato, tossico e maleodorante.
Nel 1863 fu promulgata la legge denominata "Alkali Act" che cercava di porre una regolamentazione allo smaltimento dell'acido cloridrico prodotto durante il processo Leblanc e in seguito l'invenzione del processo Solvay che permetteva di ottenere il carbonato di sodio senza scarti tossici determinarono l'abbandono del processo Leblanc su scala industriale.
Il processo Solvay per la produzione del carbonato di sodio deve il suo nome al chimico belga Ernest Solvay che lo brevettò nell'aprile del 1861.
Grazie al processo Solvay si può produrre a livello industriale sia il carbonato acido di sodio o idrogenocarbonato di sodio NaHCO3 (noto comunemente come bicarbonato di uso domestico) sia il carbonato di sodio Na2CO3 usato in altri procedimenti industriali come per esempio quello per la produzione del sapone.
Il processo solvay è usato ancora oggi, anche se in declino in quanto risulta più economico ricavare il carbonato di sodio sfruttando il "trona", che è un minerale (carbonato acido di sodio biidrato) estraibile dai laghi salati di tutto il mondo.
Nel processo Solvay si sfrutta la differenza di solubilità in acqua a 25°c tra NH4Cl (37,3 g/100 cc di soluzione) ed il NaHCO3 formato (10,0 g/100 cc) nella seguente reazione tra una soluzione di NaCl (cloruro di sodio) ed NH3 (ammoniaca) con CO2 fatta gorgogliare nella soluzione dove forma acido carbonico (H2CO3):
NaCl + NH3 + H2CO3 → NaHCO3 + NH4Cl
Questa reazione é di equilibrio, ma viene continuamente spostata a destra dalla precipitazione del sale che viene così separato per filtrazione.
La reazione in acqua del bicarbonato di sodio é leggermente basica per l’ idrolisi dello ione HCO3- ; una soluzione di 50 g/l, infatti, ha un pH = 8,5 a 25°c.
Una sua proprietà chimica importantissima, sfruttata nel processo Solvay, é che si decompone sopra i 50°C in carbonato di sodio (Na2CO3) e anidride carbonica (CO2):
2NaHCO3 → Na2CO3 + CO2 + H2O
Così il prossimo passo del processo Solvay consiste nel recuperare il NaHCO3 formato e scaldarlo decomponendolo a circa 70°C.
Il cloruro di ammonio NH4Cl della soluzione viene riciclato recuperando l’ NH3 utilizzando l'ossido di calcio (calce viva) secondo la seguente reazione:
CaO + H2O → Ca(OH)2
Ca(OH)2 + 2 NH4Cl → CaCl2 + 2 NH3 + 2 H2O
Siccome l'ammoniaca viene reimmessa nel ciclo, l'unico prodotto di scarto del processo è il cloruro di calcio CaCl2 che può essere commercializzato e utilizzato come anticongelante.
Il Na2CO3 è un sale alcalino e caustico. Una soluzione di 50 g/l a 25°C ha un pH di 11,5
La saponificazione è una reazione di idrolisi effettuata su un estere in ambiente basico. Infatti come mostrato sotto, viene fatta avvenire utilizzando come reagente una base forte come l'idrossido di sodio o di potassio.
Si parla di saponificazione quando l'estere di partenza ha un peso molecolare relativamente elevato, con una catena di almeno 12 atomi di carbonio.
Il nome saponificazione deriva dal fatto che questa reazione porta alla formazione del sapone di uso comune.
Gli esteri sono sostanze organiche formate da due radicali organici legati dal gruppo funzionale [-O-C=O-]. Nel caso della preparazione del comune sapone, si parte dai trigliceridi di acidi grassi presenti in grassi animali, come il sego bovino, o olii vegetali come l'olio di oliva.
La reazione che avviene in questo caso è descritta dallo schema seguente:
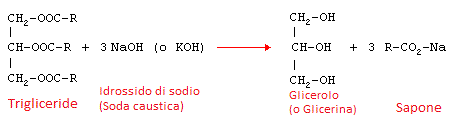
In cui vediamo a sinistra la struttura dell'estere formato dal glicerolo (detto anche glicerina) più i radicali "R" dell'acido grasso. Nel caso dell'olio di oliva questo radicale potrebbe essere quello dell'acido oleico CH3(CH2)7CH=CH(CH2)7COOH di cui l'olio di oliva è costituito per buona parte.
Il risultato della reazione è glicerolo più il sale dell'acido grasso considerato che nell'esempio, avendo usato idrossido di sodio nei reagenti (NaOH) sarà un sale di sodio.
Proseguendo la dimostrazione riportata nell'articolo a questo link, dimostriamo che l'elettore che abbiamo chiamato Mister X, oltre che essere sicuramente decisivo per una data scelta A>B, lo è per qualunque alternativa fra i candidati.
Nella prima parte della dimostrazione abbiamo visto che Mister X, che potremmo a questo punto chiamare anche "il dittatore", risulta decisivo per A su B.
Vediamo cosa succede se consideriamo l'alternativa fra A e Q, dove Q è un candidato Qualsiasi.
| X | P |
|---|---|
| A | B |
| B | Q |
| Q | A |
Già sappiamo che X è decisivo per A su B e quindi a livello globale deve essere A>B.
Per l'assioma di unanimità deve essere B>Q perchè è preferito sia da X che da P.
Per la proprietà transitiva A>B>Q implica A>Q.
Ma siccome A vince su Q nonostante P sia in disaccordo, risulta che X è decisivo nel far vincere A su Q.
Consideriamo adesso l'effetto che fa l'opinione del dittatore considerando C e un'alternativa qualsiasi che non sia A.
| X | P |
|---|---|
| C | Q |
| A | C |
| Q | A |
Abbiamo stabilito sopra che A>Q.
Per l'assioma di unanimità C>A (sono tutti d'accordo su questo, sia X che P).
Per la proprietà transitiva C>A>Q implica che C>Q.
Ancora una volta questo rispecchia il volere del dittatore e non quello di P quindi X è decisivo nel confronto fra Q e C.
Infine consideriamo il caso in cui il confronto sia fra C e A in presenza di un altro candidato Qualsiasi.
| X | P |
|---|---|
| C | Q |
| Q | A |
| A | C |
Sappiamo da sopra che C>Q.
Per l'assioma di unanimità Q>A.
Per la proprietà transitiva C>Q>A implica che C>A.
Ancora una volta C>A rispecchia la volontà del dittatore contro quella di tutti gli altri, quindi X è decisivo anche in questo caso.
Il timpano è una sottilissima membrana, tenuta in tensione dal muscolo timpanico, e capace di entrare in vibrazione, se investita dall’onda sonora proveniente dall’esterno attraverso il condotto uditivo. Grazie alle proprietà di elasticità di tale membrana, e ad un meccanismo di amplificazione costituito dalla catena degli ossicini, la sensibilità del timpano è straordinaria: è sufficiente un livello di pressione pari 0,2 miliardesimi della pressione atmosferica per attivare la sensazione sonora; a questi livelli di pressione lo spostamento della membrana timpanica è dell’ordine di 10-9 cm (un decimo circa del raggio dell’atomo di idrogeno).
Il muscolo tensore del timpano e il muscolo stapedio, che con la loro contrazione simultanea irrigidiscono la catena, costituiscono il meccanismo di regolazione del guadagno del sistema e ci permettono di percepire suoni lievi in un ambiente silenzioso e continuare a distinguere una conversazione anche in un ambiente rumoroso come potrebbe essere una discoteca.
Si deve ad Aristotele la piena comprensione dell’importanza di tre principi del nostro ragionare: il principio di identità, di non-contraddizione e del terzo escluso.
1. Il principio di identità afferma che dato A, A=A. Tale principio non è formalmente presente negli scritti aristotelici, ma da Parmenide (VI-V sec. a.C) agli stoici (III sec. a.C.) a Duns Scoto (XIII sec.) rappresenta la versione logica del fatto che, nel ragionare corretto, il significato dei termini deve mantenersi costante.
2. Il principio di non-contraddizione sostiene che, in un enunciato, non si può affermare e negare un predicato del soggetto, nello stesso tempo e nello stesso senso. Aristotele lo esprime che così: «E' impossibile che il medesimo attributo, nel medesimo tempo, appartenga e non appartenga al medesimo oggetto e nella medesima relazione» (Metafisica IV, 1005b, 19-20).
3. Il principio del terzo escluso afferma che in un sistema a due valori, Vero e Falso un enunciato è vero o è falso: una terza possibilità è esclusa. Si tratta di un principio utile per dedurre una conclusione, diciamo A, dimostrando che il suo opposto (non-A) è contraddittorio. Sono di questo tipo tutte le dimostrazioni per assurdo.
 -0
-0  )
)
